{kind=link}
甘蓝型油菜裂角相关性状的遗传与相关分析
引用本文
崔嘉成, 刘佳, 梅德圣, 李云昌, 付丽, 彭鹏飞, 王军, 胡琼. 甘蓝型油菜裂角相关性状的遗传与相关分析. 作物学报, 2013, 39(10): 1791-1798
[CUI Jia-Cheng, LIU Jia, MEI De-Sheng, LI Yun-Chang, FU Li, PENG Peng-Fei, WANG Jun, HU Qiong. Genetic and Correlation Analysis on Pod Shattering Traits inBrassica napus L. . Acta Agronomica Sinica, 2013, 39(10): 1791-1798] 
Permissions
[CUI Jia-Cheng, LIU Jia, MEI De-Sheng, LI Yun-Chang, FU Li, PENG Peng-Fei, WANG Jun, HU Qiong. Genetic and Correlation Analysis on Pod Shattering Traits in
Copyright©2013, Editorial office of Acta Agronomica Sinica
This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
甘蓝型油菜裂角相关性状的遗传与相关分析
摘要
抗裂角性是非常重要的油菜性状, 但相关研究报道较少。本研究对11个甘蓝型油菜骨干亲本品系及由其配制的30个不完全双列杂交组合在2个环境下的抗裂角指数及其他7个角果相关性状进行了遗传分析。结果表明, 抗裂角指数遗传变异显著, 遗传上受少数主效基因控制, 效应以加性为主, 显性效应和环境效应影响较小。大部分杂交组合的抗裂角指数杂种优势不显著。抗裂角指数与角果长、果皮重、千粒重和种子直径呈极显著正相关, 与结角密度和角粒数呈极显著负相关。抗裂角性相关性状中, 角果长、千粒重、结角密度和种子直径变异主要由加性方差解释; 果皮重和每角粒数主要由显性方差解释。亲本评价分析指出, 作为波里马细胞质雄性不育系统保持系的ZS11B和恢复系的R11其抗裂角性的一般配合力高, 是培育抗裂角杂交油菜品种的首选直接亲本。
关键词:
油菜; 遗传效应; 抗裂角指数; 相关分析
Genetic and Correlation Analysis on Pod Shattering Traits inBrassica napus L.
Abstract
Pod shattering resistance is a very important trait of oilseed rape, and the genetic research has been extremely weak. In this study, pod shattering resistance index (SRI) and other seven pod traits were analyzed based on data collected from a 6×5 incomplete diallel design at two environments. The results demonstrated that obvious genetic variation existed among elite breeding lines and their crosses for pod SRI. Pod shattering resistance was likely controlled by few genes with mainly additive effects, and dominant effects were much less important, whereas the environments played only minor role for pod SRI. There was no significant heterosis in most of the crosses. Pod SRI was detected to be significantly positively correlated with pod length, pod wall weight, 1000-seed weight and seed diameter, but negatively correlated with pod density and number of seeds per pod. The genetic variations of pod length, 1000-seed weight, pod density and seed diameter were mainly explained by additive effects, whereas those of pod wall weight and number of seeds per pod were mainly affected by dominant effects. Among the 11 elite breeding lines, ZS11B and R11 were recommended for using as direct parental lines for making pod shattering resistant hybrid varieties as maintainer and restorer lines for Polima cytoplasmic male sterility system, respectively, due to their higher general combining ability on pod SRI and other yield component traits.
Keyword:
Oilseed rape; Genetic effect; Pod shattering resistance index; Correlation analysis
引言
裂角是油菜在长期进化过程中形成的与繁衍生息相关的生物学特性[1]。但是在生产实践中, 裂角在油菜成熟和收获时造成严重的产量损失[2]。目前油菜栽培种中抗裂角性最强的是芥菜型油菜, 其次是白菜型油菜, 最差的是甘蓝型油菜[3]。研究表明, 油菜成熟后因角果炸裂导致10%以上的产量损失[4], 收获延迟会导致近20%的产量下降[5], 而在不良 气候条件下特别是机械收获时产量损失最高达 50%[6,7]。随着我国农业现代化发展, 油菜生产机械化是发展油菜生产、提高油菜生产效益的唯一出路。因此, 控制油菜收获时的裂角落粒是当前油菜科研中非常重要的课题。
控制油菜裂角落粒通常有3种途径。一是分段收获, 在油菜籽粒没有完全成熟时将油菜割倒, 待植株完全脱水后再机械捡拾脱粒。由于收割时没有完全成熟, 角果含水量大, 减少了角果的炸裂和落粒。但是产量潜力没有充分发挥, 存在很多绿籽和红籽, 籽粒中叶绿素含量偏高, 直接影响菜籽油的品质。二是喷洒化学药剂增加角果的抗裂性。Szot等[8]研究发现喷施SPODNAM可减少10%~45%的产量损失, 但污染环境, 且在油菜成熟期操作困难, 增加生产成本。同时这种化学药剂减少角果炸裂的原因是在角果表面形成保护膜, 由于收获时菜籽的含水量较高, 该保护膜不利于菜籽水分的蒸发, 影响种子的干燥和贮藏[9]。三是通过种质资源的创新如种间杂交[3]、人工合成[10]等方法不断引进抗裂角基因资源, 培育抗裂角甘蓝型油菜品种。在我国冬油菜主产区长江流域, 油菜收获时常常遇到多雨潮湿天气, 为机械操作带来不便, 所以前2种减少落粒的途径均不适用。只有通过挖掘现有油菜品种及种质资源中的抗裂角材料, 培育抗裂角性强的油菜新品种, 延长油菜品种的适宜收获期, 减少油菜充分成熟后机械收获造成的落粒损失, 才能从根本上解决油菜机械化生产的问题。
由于油菜抗裂角性普遍较差, 大多数油菜品种不抗裂角, 少数抗裂角较强的材料多为种间杂交或人工合成甘蓝型油菜的后代[3,11,12], 有的还是染色体附加系[13], 关于油菜裂角性的遗传研究也就相对缺乏。Morgan等[14]通过利用抗裂角性强的人工合成甘蓝型油菜衍生品系与抗裂角性差的油菜品系进行双列杂交, 研究了油菜抗裂角性的遗传及其与角果性状的关系, 认为抗裂角性主要受加性效应控制, 受显性效应影响较小; 抗裂角性与果皮厚度呈正相关, 与角果密度、角果长度、角果宽度和角粒数不相关, 与果喙长度显著负相关。文雁成等[15]对229份油菜品种(系)的鉴定分析认为, 抗裂角性与角果密度呈极显著负相关, 与角果长度、宽度、喙长、果皮厚度及角粒数均呈极显著正相关。上述有限的研究由于所用材料的不同, 结果相差较大, 也没有考虑到环境因素对遗传效应的影响。近年来随着裂角性鉴定方法的改进和生产要求的急迫, 国内利用改进后的随机碰撞法对我国现有油菜品种(系)进行了抗裂角性鉴定, 发现我国甘蓝型油菜品种间抗裂角性存在明显差异[4,15], 为研究油菜裂角性遗传及油菜品种的抗裂角性遗传改良奠定了基础。本研究采用近年来筛选出的具有不同裂角抗性的油菜杂交种骨干亲本, 配制不完全双列杂交组合, 在不同环境中鉴定亲本和杂种F1代的抗裂角指数及角果相关性状, 利用加性-显性与环境互作的遗传模型[16]分析角果相关性状的遗传效应及其与抗裂角性的相关性, 为油菜抗裂角性遗传改良和杂种优势利用提供了理论依据, 为从现有的材料中鉴定出有利于提高抗裂角性的基因型资源奠定了物质基础。
1 材料与方法
1.1 试验材料
以11个甘蓝型油菜育种轮回选择群体中材料为亲本, 分别为0028、1019B、1055B、6098B、8908B、ZS11B (中双11选系)(波里马细胞质雄性不育系的保持系)和HPR1、R2、R6、R9、R11 (恢复系), 均为中国农业科学院油料作物研究所育成品种(系), 其中ZS11B为油菜生物技术育种课题组刘贵华研究员惠赠, 其余材料为本课题组选育的骨干亲本。以6个保持系为母本, 5个恢复系为父本, 采用不完全双列杂交设计配制杂交组合, 获得30个F1杂种, 加上11个亲本, 共取得41份材料的性状鉴定数据。
1.2 试验方法
于2011年10月至2012年5月分别在湖北省的新洲(E1)和汉川(E2)进行田间试验。完全随机区组设计, 3次重复, 4行区, 行长2 m, 行距0.33 m, 每行留苗18株。成熟期每小区收取10株, 在相同条件下自然干燥一个月后考种, 取10株平均值得到角果密度, 并在每个单株的主花序上、中、下部随机取20个发育正常的角果, 每小区总计200个角果, 用保鲜袋装好备用。
鉴定抗裂角性采用优化后的随机碰撞法。随机选取20个角果烘干, 放入直径14.8 cm、高7.4 cm的圆柱型塑料容器内, 容器中放置8个直径为14 mm的钢球, 将容器放在280 转 min-1、振幅为24 mm的摇床上(HQ45Z, 武汉中科科仪技术发展有限公司)振荡10 min, 每2 min记录一次破裂角果数, 每份材料重复3次, 计算抗裂角指数。裂角指数= ∑ xi×(6- i)/100(角果数×总次数), 式中, xi 为第 i次破损的角果数, 1≤ i≤5; 抗裂角指数=1-裂角指数。每份材料选取100个角果测定角果长度、果皮重量、果喙长和每角粒数, 利用万深SC-I型种子大米自动数粒分析仪系统测定种子直径和粒重, 计算千粒重。
1.3 数据分析
运用加性-显性与环境互作的遗传模型分析数据[16], 利用QGAStation分析软件(http://ibi.zju.edu.cn/software/ qga/)估算各项遗传参数, 采用以基因型为抽样单位的Jackknife抽样方法估算各项统计量的标准误差, 然后检验显著性。
应用SAS软件进行抗裂角指数的方差分析和计算变异系数及平均值。
2 结果与分析
2.1 抗裂角指数的性状表现
41份材料中, 2个F1杂种在汉川试验点因出苗不齐而缺失数据, 所以参与分析的数据来自79个试验小区。方差分析表明, 试验材料间存在极显著变异( P<0.0001), 而区组间、试验地点和材料×地点等变异项不显著(表1)。说明抗裂角性状的变异主要受遗传因素影响, 同时也说明优化后随机碰撞法鉴定的抗裂角指数的稳定性和重现性好。79个小区间抗裂角指数变异幅度为0~0.47, 平均值为0.1128, 变异系数为82.95%, 说明试验所用材料的抗裂角性遗传差异明显, 意味着有可能从这些材料中鉴定出有利于提高抗裂角性的基因型资源。
各亲本在新洲和汉川两地差别不大, 趋势一致。在新洲各亲本平均抗裂角指数最小值为0, 最大值为0.4300, 平均值为0.1382; 汉川的平均抗裂角指数最小值为0, 最大值为0.4700, 平均值为0.1464 (图1)。相关分析表明各亲本在两地的抗裂角指数之间相关系数为0.9121 ( P<0.0001), 呈极显著正相关, 也说明亲本抗裂角性受环境影响小。若将抗裂角指数在0.3以上和0.1以下分别划分为相对抗裂角和相对易裂角类型, 则1055B和ZS11B为抗裂角亲本, 1019B和R11为较抗裂角亲本, 其余为不抗裂角 亲本。
| 表1 抗裂角指数的方差分析 Table 1 ANOVA of pod shatter resistance index |
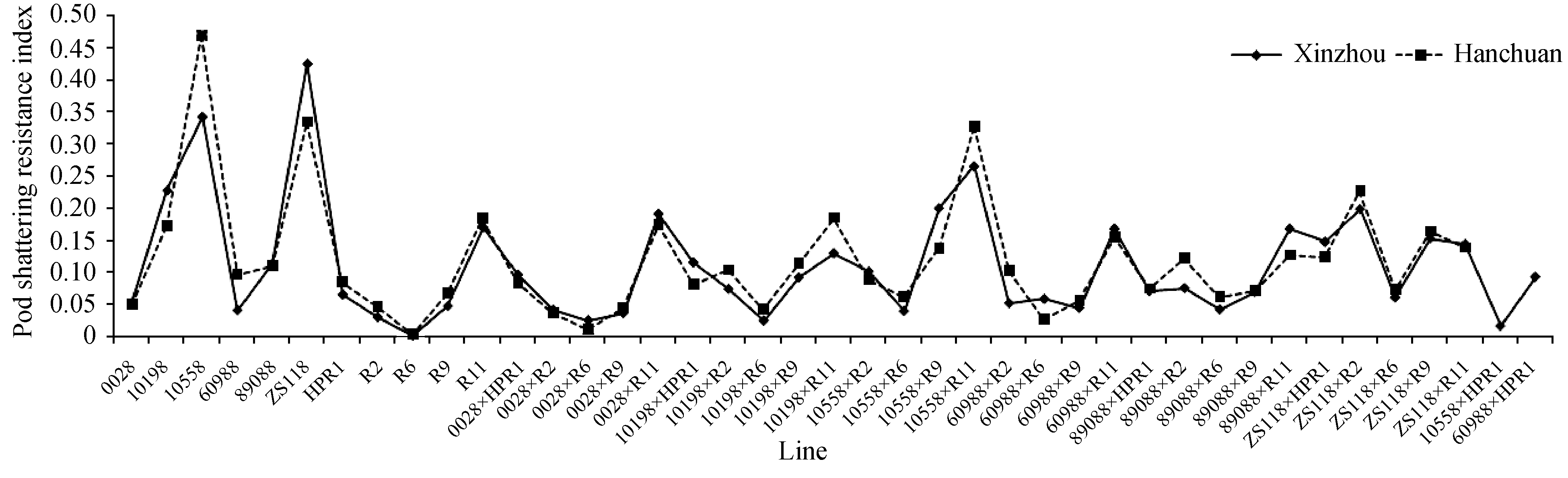 | 图1 11个亲本及30个杂交组合的抗裂角指数Fig. 1 Pod shatter resistance index of 11 parents and 30 crosses |
杂种F1代的抗裂角指数间也存在很大变异, 在新洲F1代抗裂角指数变异范围为0.02~0.27, 平均值为0.10; 而在汉川抗裂角指数变异范围0.01~0.33, 平均值为0.11; 平均值、最大值和变异范围均小于亲本间。两地间杂种F1代抗裂角指数的差别仍然较小, 相关系数为0.8975 ( P<0.0001), 相关性也是极显著。杂种F1代中只有少数组合表现超亲优势, 大部分组合都表现无优势或中亲优势。抗裂角性最高的组合为1055B×R11, 在新洲和汉川分别为0.27和0.33。抗裂角性较高的组合还有0028×R11、1019B× R11、6098B×R11、8908B×R11、ZS11B×R2和ZS11B×R9等, 在两地的抗裂角指数都在0.1以上。
2.2 抗裂角指数与角果性状的遗传相关分析
为了研究抗裂角性与角果性状间的相关性, 本试验同时鉴定了与抗裂角可能相关的其他角果性 状, 包括角果长度、果喙长、果皮重、每角粒数、结角密度、千粒重和种子直径。采用QGAStation软件作遗传相关分析, 将抗裂角指数与7个角果相关性状遗传相关分解为 rA (加性效应相关)、 rD (显性效应相关)、 rP (表型相关)和 rG (基因型相关) 4个相关指标。表型与基因型相关分析结果表明, 抗裂角指数与角果长、果皮重、种籽直径和千粒重均呈极显著正相关, 与每角粒数和结角密度则呈极显著负相关, 与果喙长相关性不明显(表2)。在所有显著性相关的性状中, 角果长和果皮重与抗裂角指数的相关系数最高, 基因型相关系数在0.45以上。与抗裂角指数的基因型相关系数绝对值由大到小的性状依次是果皮重>角果长>种子直径>结角密度>千粒重>每角粒数>果喙长。所有性状与抗裂角指数的表型相关系数和基因型相关系数之间差异较小, 也验证了这些性状主要是受遗传因素影响, 环境作用较小。在基因型相关中, 以加性遗传相关最为明显, 其中抗裂角指数与果皮重的加性遗传相关系数高达1, 与角果长的加性遗传相关系数也达到0.606。显性遗传相关较小, 且多不显著, 只有角果长和结角密度与抗裂角指数的显性遗传负相关达到极显著水平, 但相关系数都很小, 绝对值都在0.046以下。
2.3 抗裂角相关性状的遗传方差组成
分析油菜角果相关性状的各项遗传方差分量占表型方差的比率(表3)表明, 各性状的加性方差比率均达到极显著水平, 以角果长、结角密度和抗裂角指数的加性方差比率最高, 均达到50%以上。有4个性状即果喙长、千粒重、结角密度和种子直径的加性×环境互作方差比率达到极显著水平。除千粒重外, 所有性状的显性方差比率都达到显著或极显著水平, 以果喙长和每角粒数的显性方差比率最高, 达40%以上; 抗裂角指数、果皮重、千粒重、结角密度和种子直径的显性×环境互作方差比率也达到极显著水平。
| 表2 抗裂角指数与角果性状间相关系数 Table 2 Correlation coefficient between pod SRI and other pod traits |
| 表3 油菜角果相关性状方差分量比率估算值 Table 3 Estimated proportions of variance components for pod traits |
除果喙长、果皮重和每角粒数外, 其余性状的加性方差比率均高于显性方差比率。除抗裂角指数、千粒重、结角密度和种子直径外, 其余性状的显性方差占比均高于显性×环境互作方差比率(表3)。除千粒重和种子直径外, 其他性状的遗传效应比率( Va/ Vp+ Vd/ Vp)都在55%以上, 其中以角果长最高, 达87.2%, 抗裂角指数的遗传效应比率也接近80%。遗传力分析结果也表明抗裂角指数与角果长、果喙长、每角粒数和结角密度一样, 都属于遗传力较高的性状, 广义遗传力均在60%以上。说明角果相关性状主要受遗传因素影响, 包括加性和显性效应, 同时受环境影响较小。
2.4 亲本抗裂角指数的加性效应
根据遗传方差分析结果, 角果性状受加性效应影响明显, 且所有性状的加性效应明显高于加性×环境互作效应, 因此对各亲本的加性主效应进行了具体分析。
11个亲本中, 抗裂角指数的加性效应达极显著的8个, 达显著的2个。其中4个为正值的全部为极显著, 另外6个为极显著或显著的负值。具有正加性效应的亲本中, 以1055B最高, 达0.101, 其后依次为ZS11B、R11和1019B, 加性效应值分别为0.081、0.034和0.020; 具有负加性效应的亲本中, 绝对值最高的为R6 (0.083), 其后依次为R2、HPR1、R9、0028和6098B, 加性效应的绝对值变幅为0.017~0.042。除R9和6098B的负加性效应为显著外, 其余均为极显著, 仅8908B为不显著的负效应。
分析与抗裂角指数显著相关的6个角果性状(果喙长与抗裂角指数的相关性不显著, 不在一起分析)在抗裂角指数具有正向加性效应的亲本中的遗传效应, 发现在这4个亲本中, ZS11B所有考察性状的加性效应值均为正值, 除了每角粒数和结角密度不显著外, 角果长、果喙长、果皮重、千粒重和种籽直径的加性效应均达到极显著水平。抗裂角指数加性效应值最大的1055B, 仅果皮重的加性效应值为极显著的正值, 角果长和种籽直径的加性效应值不显著, 果喙长、每角粒数、千粒重和结角密度的加性效应均为极显著的负值。另外2个抗裂角指数的加性效应为显著正向效应的亲本中, R11角果长、果皮重、千粒重、结角密度和种籽直径等性状的加性效应值也都是正向的, 1019B每角粒数的加性效应也为正向极显著。
2.5 抗裂角指数的显性效应与杂种优势
根据遗传方差分析结果, 虽然抗裂角指数主要受加性效应控制, 但是显性方差比率和显性×环境互作方差比率均达到了极显著水平, 且显性×环境互作方差比率大于显性方差比率。因此在分析显性效应时, 需考虑显性×环境互作效应的影响。由于抗裂角指数的显性方差和显性×环境方差比率均较小, 在20%以下, 因此在所有30个杂交组合中, 仅有7个组合表现出显著的显性或显性与环境互作效应, 绝对值变幅为0.017~0.086 (表5)。其中4个组合(0028×R11、ZS11B×R2、0028×HPR1和1055B×R9)的抗裂角指数均表现出正向显性主效应, 以1055B×R9显性主效应值最高。2个组合表现极显著的负向显性主效应, 另外2个组合负向显性主效应不明显。有4个组合表现出显著的显性×环境互作效应, 但组合间显性×环境互作效应在两试验点表现不同。组合ZS11B×R2在汉川表现正向显性×环境互作效应, 组合0028×HPR1在新洲表现正向显性×环境互作效应, 其余地点的显性×环境互作效应均不显著。组合ZS11B×R11和1019B×R11分别在汉川和新洲表现出显著负向的显性×环境互作效应。
| 表4 亲本各性状加性效应预测值 Table 4 Predicted additive effects of different pod traits for parent lines |
| 表5 部分F1杂交组合抗裂角指数的显性效应预测值 Table 5 Predicted dominance effects of pod SRI for some F1 crosses |
杂种的基因型预测值度量的是杂种在不同环境中不受环境因素干扰的平均表现[17]。本试验中所研究的杂交组合的基因型值都达到了显著或极显著水平, 说明基因型对杂种F1代抗裂角指数的表现起决定作用。30个组合平均基因型值为0.104, 变幅为0.004~0.293; 群体平均优势平均值为-0.187, 变幅为-1.060~0.480; 群体超亲优势平均值为-0.873, 变幅为-2.880~0.108。仅4个组合表现出显著的基因型×环境互作效应, 其中正向和负向各占2个, 正向的组合为0028×HPR1和ZS11B×R2。仅1个组合表现极显著的正向群体平均优势, 1个组合表现显著的负向平均优势。群体平均优势×环境的互作不明显, 仅1个组合(1019B×R11)表现出显著的负向的互作效应。除1个组合(0028×HPR1)表现显著的正向超亲优势外, 5个组合表现极显著的负向超亲优势。这6个具有超亲优势的组合中, 有4个的表现受到环境的显著影响。
3 讨论
油菜抗裂角性在机械化生产中的重要性日益突出, 但由于抗裂角遗传变异来源复杂, 抗裂角鉴定方法不统一, 关于油菜抗裂角性遗传的研究结果难以整合比较。Mongkolporn等[18]发现白菜型油菜角果的抗裂角性由2个主效隐性基因控制。彭鹏飞[19]利用油菜F2群体定位初步获得2个主效抗裂角QTL。而文雁成[20]认为油菜抗裂角性是受微效基因控制的数量性状, 并分别在郑州和武汉定位到9个和4个抗裂角性状的QTL, 对表型的总贡献率分别达到45.9%和38.6%。最近, Hu等[21]研究认为油菜角果的抗裂角性变异受少数基因控制, 并检测到1个主效QTL, 对表型的贡献率高达47%。本研究利用不完全双列杂交2个地点的试验数据, 运用混合线性模型对抗裂角性的遗传效应及其与环境互作的分析, 发现油菜的抗裂角性主要受遗传因素的作用, 受环境的影响较小, 且起主要作用的加性效应与环境的互作为零, 总体上抗裂角指数的杂种优势不显著, 因此推断至少对于本试验群体, 抗裂角性的遗传属于少数主效基因控制的性状, 以加性效应为主, 受显性效应影响较小。这一研究结果与Morgan等[14]利用人工合成的抗裂角甘蓝型油菜品系的双列杂交研究结果一致, 说明在不同来源的抗裂角性资源中, 有可能有相似或相同的遗传模式。
了解性状间的遗传相关对于多个性状同步改良、性状的间接选择和打断决定性状的基因位点间的连锁累赘均有重要意义[22]。何余堂等[23]、Morgan等[2]、Wang等[3]和文雁成等[15]对油菜抗裂角性与角果相关性状关系的研究, 都基于简单的表现型相关, 没有涉及到遗传相关。本研究将抗裂角指数与角果相关性状间的相关分解为加性相关和显性相关, 并分析了基因型相关和表型相关, 表明抗裂角指数与角果长、果皮重、籽粒大小呈极显著正相关, 与结角密度、角粒数呈显著负相关, 而与果喙长的相关性不明显。本研究得出的抗裂角指数与角果长正相关的结论, 与文雁成等[15]的结果相同, 但与Wang等[3]结论相反。由于一般情况下果喙长与角果长呈显著正相关( r=0.519), 通常认为果喙长与角果长一样, 与抗裂角性呈正相关[15], 但本研究得出果喙长并不像角果长一样与抗裂角指数呈显著正相关, 而是相关性不明显, 验证了Wang等[3]得出的抗裂角性与果喙长不相关的结论。抗裂角指数与果皮重呈极显著正相关, 与文雁成等[15]和Morgan等[2]研究结果一致。与每角粒数和结角密度呈极显著负相关, 与何余堂等[23]研究结果类似。但本试验结果发现抗裂角指数与千粒重和种子直径极显著正相关, 则与何余堂等[23]的结果不同, 可能与本试验选用的3个抗裂角性稍强的亲本中有两个是大粒材料有关。但在育种实践中, 我们观察到大粒材料往往果皮较厚, 田间抗裂角性表现也较好。本研究结果和国内外其他研究团队结果间的差异可能与所用材料的遗传背景、群体大小和抗裂角性鉴定方法有关。但总体看来, 大多数的研究结果支持抗裂角指数与角果长度和果皮厚度呈显著正相关的结论。
性状间加性相关系数决定是否可在早代对某一个或几个性状进行间接选择[24]。本研究与抗裂角指数加性正相关极显著且相关系数高的为果皮重和角果长。尽管角果长与果皮重均与抗裂角指数呈显著正相关, 但果皮重与抗裂角指数的加性相关系数比角果长与抗裂角指数的加性相关系数更高, 说明果皮重并不仅仅是通过角果长来影响抗裂角性, 果皮厚度的作用也不可小视。育种实践中应该通过对长角果和厚角果皮的定向选择来间接提高抗裂角性, 不仅直观性强易于在田间选择时操作, 同时由于长角果和厚角果皮多数情况下还与产量组成性状如粒重和粒数呈正相关, 还有利于产量的提高。
本研究所用11个亲本中, 抗裂角指数正向加性效应值(一般配合力)较大的前2个亲本是保持系群体中的1055B和ZS11B。但是由于ZS11B所有考察性状的加性效应值均为正值, 且大部分性状的加性效应值均达到极显著水平。同时, ZS11B所组配的杂交组合抗裂角指数在不同的环境条件下更稳定, 如组合ZS11B×R2和ZS11B×R11在不同地点显性效应与环境互作值方向一致。而抗裂角指数加性效应值最大的1055B, 除果皮重的加性效应值为极显著的正值外, 其余性状的加性效应值或为不显著的正值, 或为极显著的负值。因此, 在培育抗裂角品种时, ZS11B是组配杂交种时保持系直接亲本的首选。在恢复系群体中, R11的加性效应值最高, 也是恢复系群体中唯一具有正向加性效应的亲本, 是组配杂交种时直接亲本(恢复系)的首选。
4 结论
证实抗裂角指数存在明显的遗传变异, 具有较大的改良潜力。角果抗裂角性的遗传以加性效应为主, 受显性效应影响较小, 可以在早代对此性状选择。角果长、果皮重、千粒重和种子直径与抗裂角指数呈极显著正相关; 每角粒数和结角密度与抗裂角指数呈极显著负相关; 果喙长与抗裂角指数间不存在相关性。在培育抗裂角品种时, ZS11B是组配杂交组合首选的保持系直接亲本, R11是组配杂交组合首选的恢复系直接亲本。
致谢: 感谢浙江大学朱军教授举办的数量遗传分析及QTL定位研讨班并无偿提供QGAstation数据分析软件; 感谢安红玉、桑世飞、康中德对性状鉴定的贡献。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|